

Trong y văn, các nghiên cứu về trường thọ (longevity) đã và đang gặt hái được nhiều kết quả. Quá trình lão hóa khi được nhìn nhận dưới góc độ sinh học phân tử giúp hé lộ nhiều bí ẩn của tự nhiên. Mặc dù trường thọ liên quan đến sự cân bằng nội môi (homeostasis) trong dòng chảy thông tin từ gene cho tới protein, chỉ hai đầu cực của dòng chảy là được nghiên cứu kỹ lưỡng. Đoạn giữa của dòng chảy, tức các quá trình xảy ra ở RNA, bao gồm cắt tiền RNA thông tin (pre-mRNA splicing), vẫn chưa được khám phá rõ ràng.
Trong công trình xuất bản trên tạp chí Nature ngày 5.12.2016, các nhà khoa học tại Đại Học Harvard, dưới sự hướng dẫn của giáo sư trợ lý (assistant professor) William Mair, mở ra một xa lộ mới về pre-mRNA splicing cho các nghiên cứu về trường thọ trên loài giun tròn (roundworm) Caenorhabditis elegans).
Để có thể quan sát được những sự thay đổi về kiểu cắt (splicing pattern), Caroline Heintz, tác giả số một của bài báo, và cộng sự của Cô sử dụng gene báo cáo cắt huỳnh quang nội bào (in vivo fluorescent splicing reporter) ret-1 (Hình 1a).
 |
Hình 1a: Sơ đồ gene ret-1 splicing reporter.
Hình 1b: Sự biểu hiện của GFP và mCherry ở ngày 1 do sự cắt đặt hiệu của ret-1.
ret-1 minigene reporter sử dụng eft-3 promoter, và điểm đặt biệt của nó nằm ở chỗ: nếu pre-mRNA của ret-1 gene không bị cắt exon 5 (exon inclusion), gene mã hóa protein huỳnh quang xanh (GFP) sẽ được biểu hiện (màu xanh); và nếu pre-mRNA của ret-1 gene bị cắt exon 5 (exon skipping), gene mCherry được biểu hiện (màu đỏ). Loài giun C. elegans có lớp da trong suốt giúp dễ dàng quan sát được sự biểu hiện màu này. Thêm vào đó, việc bị cắt hay không bị cắt exon còn có tính địa phương. Cụ thể, exon inclusion thường xảy ra ở các tế bào đường ruột (intestine), trong khi exon skipping xảy ra ở hạ bì (hypodermis), thành cơ tế bào (body muscle wall), và hệ thần kinh trung ương (Hình 1b).
Tiếp đó, các nhà khoa học kiểm tra xem gene báo cáo ret-1 có đáp ứng với sự gián đoạn thể cắt (spliceosome disruption) hay không. Trong spliceosome của giun C. elegans, hrp-2 là một thành phần cốt lõi. Khi ức chế hrp-2 bằng giao thoa RNA (RNA inteference – RNAi), giải trình tự RNA cho thấy sự cắt tiền mRNA của gene ret-1 bị thay đổi.
Sau đó, các nhà khoa học sử dụng gene báo cáo để theo dõi hoạt động cắt ở giun đang lão hóa. Kết quả cho thấy, khi giun ở giai đoạn ấu trùng (larva) tới trước ngày số 5, mẫu cắt tuân theo quy luật nêu ở trên (Hình 1b). Tuy nhiên, vào ngày số 5, mặc dù kiểu hình của các con giun là đồng nhất (homogenous), kiểu cắt của chúng lại không đồng nhất (heterogenous) (Hình 1c). Sự khác nhau này biểu hiện ở cả hai mức độ, mức độ quần thể (giữa các con giun với nhau) và mức độ cá thể (giữa từng tế bào trong cùng một con giun). Sang ngày thứ 7, tất cả các con giun đều thay đổi mẫu cắt. (Hình 1d)

Hình 1c và d: Kiểu cắt (splicing pattern) đối với tiền RNA thông tin (pre-mRNA) ở giun C. elegans. Màu xanh (GFP) cho thấy exon 5 được giữ lại (exon inclusion), và màu đỏ (mCherry) cho thấy exon 5 bị loại bỏ (exon skipping). Ở ngày 1, kiểu cắt được xem là “trẻ trung” (youthful). Sang ngày 5, kiểu cắt đã thay đổi từ exon inclusion sang exon skipping. Tới ngày 7 thì tất cả các tế bào hầu như tuân theo kiểu cắt exon skipping. Bảng bên dưới tóm tắt lại thông tin một cách ngắn gọn hơn.

Như vậy, quá trình lão hóa làm thay đổi kiểu cắt tiền RNA thông tin ở tế bào, và tỉ lệ thay đổi này khác nhau giữa các cá thể.
Tiếp theo, các nhà khoa học xem xét vai trò của việc giới hạn chế độ ăn (dietary restriction – DR) lên sự cắt ret-1. Các nghiên cứu trước cho thấy, khi giun C. elegans được nuôi trong điều kiện DR, tuổi thọ của chúng kéo dài hơn 65%. Heintz và cộng sự quan sát hai nhóm giun, một nhóm nuôi trong điều kiện giới hạn chế độ ăn (nhóm DR) và một nhóm nuôi trong điều kiện đầy đủ thức ăn (nhóm ad libitum hay AL). Kết quả (tóm tắt trong bảng dưới) cho thấy có sự khác nhau về kiểu cắt ở cả hai mức độ, quần thể (population) và cá thể (individual).

Mặc dù các con giun ở nhóm AL đã được chọn lựa để có kiểu gene tương đồng (isogenic), sự khác nhau về tỉ lệ thay đổi kiểu cắt giữa các giun lớn đáng kể (bất đồng nhất quần thể – population heterogeneity). Vậy lý do của điều này là do đâu? Các nhà khoa học chọn các con giun ở nhóm AL có cùng ngày tuổi (ngày thứ 6) và chia thành hai nhóm dựa trên kiểu cắt để quan sát.
Nhóm A: có kiểu cắt exon 5 skipping, giống như kiểu cắt của giun ở nhóm AL.
Nhóm B: có kiểu cắt exon 5 inclusion, giống như kiểu cắt của giun ở nhóm DR.
Tuổi thọ trung vị (median) của nhóm B cao hơn hẳn nhóm A, vốn là nhóm có các biểu hiện sớm của sự suy giảm điều tiết kiểu cắt (splicing deregulation). Như vậy, sự khác nhau về tỉ lệ thay đổi kiểu cắt ở giun nhóm AL có liên quan tới sự khác nhau trong tiến trình lão hóa kiểu dại. Hơn nữa, hiệu suất cắt (splicing efficiency) giảm dần ở nhóm AL hơn là ở nhóm DR, và kiểu cắt có thể được xem như là một chỉ thị tiên đoán (predictor) cho kỳ vọng sống sau sinh (life expectancy).
Tiếp theo, các nhà khoa học nghiên cứu xem các yếu tố cắt (splicing factor) có ảnh hưởng gì lên hiệu ứng trường thọ của chế độ ăn giới hạn (DR) hay không. Lần này, Heintz và cộng sự sử dụng giun bị đột biến eat-2(ad1116) để có nhóm DR thay vì nuôi giun trong điều kiện DR. Như vậy, có hai cách để thu được nhóm DR: 1) các giun dạng dại (wild type) được nuôi trong điều kiện DR, và 2) các giun có đột biến eat-2(ad1116) (gọi là mô hình di truyền – genetic model).
Các nhà khoa học tiến hành sàng lọc di truyền ngược có mục đích (targeted reverse genetic screening) đối với các thành phần trong thể cắt (spliceosome). Các yếu tố cắt này gồm cả các thành phần chủ chốt của spliceosome lẫn các protein bám RNA thuộc hai họ protein SR và hnRNP. Kết quả sàng lọc cho thấy, yếu tố cắt protein bám điểm nhánh SFA-1 (branch-point binding protein splicing factor 1) khi bị ức chế bằng RNAi thì giun ở nhóm DR hoàn toàn không được kéo dài tuổi thọ, còn giun ở nhóm AL thì không bị giảm tuổi thọ.
Khi ức chế gene sfa-1 bằng RNAi, các nhà khoa học nhận thấy 1) biểu hiện gene sfa-1 giảm 50%, 2) không ảnh hưởng tới biểu hiện gene uaf-1 vốn nằm trên cùng một operon với sfa-1, 3) không ảnh hưởng tới tỉ lệ ăn (feeding rate) hay tỉ lệ bơm (pumping rate) ở giun, và 4) hiệu ứng trường thọ của DR đối với kiểu cắt ret-1 bị ức chế. Hơn nữa, khi xem xét kiểu cắt tiền RNA thông tin ở gene tos-1, chế độ ăn DR cũng làm ức chế sự thay đổi kiểu cắt của tos-1 và có sự phụ thuộc vào hoạt động của SFA-1.
Lưu ý rằng, ở đây các nhà khoa học có hai cách để biết sự thay đổi kiểu cắt: 1) sử dụng gene báo cáo cắt nội bào (in vivo splicing reporter) (trường hợp gene ret-1) và 2) phân tích PCR nếu không có gene báo cáo (trường hợp gene tos-1).
Để phân tích không thiên vị vai trò của lão hóa, chế độ ăn giới hạn, và SFA-1 lên tiến trình RNA (RNA sequencing) và biểu hiện gene, các nhà khoa học tiếp tục nghiên cứu trên nhóm AL (sử dụng dòng giun N2)và nhóm DR (sử dụng dòng giun eat-2(ad1116), DA1116). Các mẫu thu vào các ngày thứ 3, 15, và 27 (ngày 27 đối với các giun DR, giun AL chết trước ngày 27), xem bảng bên dưới.

Kết quả cho thấy, quá trình lão hóa không ảnh hưởng tới biểu hiện gene của splicing factor, nhưng có gây cảm ứng sự gián đoạn thể cắt. Các mẫu thu vào ngày 15 ở nhóm AL cho thấy có sự tăng cắt nhiễu (splicing noise). Trong khi đó, ở nhóm DR, cắt nhiễu giảm cũng như tính bất đồng nhất kiểu cắt (splicing heterogeneity). Khi ức chế sfa-1 bằng RNAi, vai trò này của DR mất đi. Như vậy, DR có vai trò chống lại các tiến trình RNA sai lệch bên trong tế bào theo tuổi, và chức năng bảo vệ này của DR có liên quan đến SFA-1.
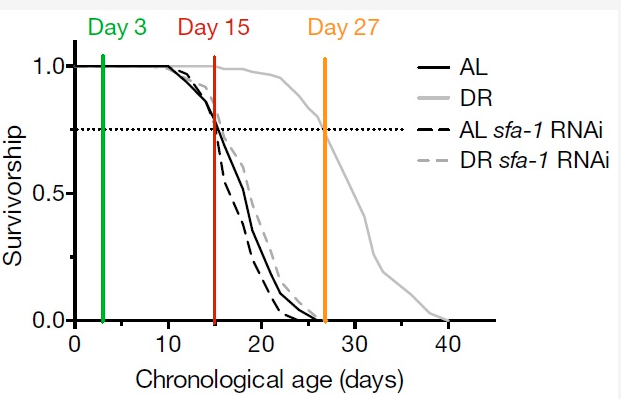
Hình 2: Minh họa mối liên hệ giữa ngày tuổi và tỉ lệ sống sót của 4 nhóm giun C. elegans: nhóm AL, nhóm DR, nhóm AL bị ức chế sfa-1, và nhóm DR bị ức chế sfa-1 bằng RNAi. Giun ở nhóm DR sống tới ngày 40 so với giun các nhóm khác.
Các nghiên cứu khác cũng cho thấy SFA-1 có vai trò trong tính mềm dẻo chuyển hóa (metabolic plasticity) (làm tăng dung tích hô hấp cực đại – maximal respiratory capacity) và chức năng ti thể ở chế độ ăn giới hạn.
Các nhà khoa học tiến hành một chuỗi các thí nghiệm nhằm tìm ra cơ chế của SFA-1 lên hiệu ứng kéo dài tuổi thọ của chế độ ăn giới hạn. Đầu tiên, các nhà khoa học để gene báo cáo ret-1 vào các giun có các đột biến tắt (null mutation) ở các gene có liên quan tới việc tăng tuổi thọ do chế độ ăn DR, bao gồm:
aak-2: mã hóa đơn vị dưới xúc tác của AMPK (cảm biến năng lượng – energy sensor)
daf-16: yếu tố phiên mã đầu nĩa (forkhead transcriptional factor)
raga-1: mã hóa Rag GTPase kết nối giữa việc thăm dò amino acid (amino acid sensing) với mục tiêu cơ học của hoạt động phức hợp 1 rapamycin (mTORC1).
Kết quả cho thấy, ở giun thiếu RAGA-1, 1) hiệu ứng của DR lên mẫu cắt ret-1 bị mất, 2) quá trình phosphorylation của đích mTORC1 RSKS-1/S6 kinase bị giảm, và 3) hiệu ứng của tuổi thọ lên SFA-1 bị ức chế. Như vậy, RAGA-1 là một mắc xích then chốt giữa DR và sự điều hòa cân bằng nội môi mẫu cắt tiền RNA thông tin.
Các nghiên cứu cho thấy việc ức chế trực tiếp mTORC1 làm tăng tuổi thọ, và mTORC1 có vai trò như một chất điều tiết (mediator) trong các hiệu ứng có lợi của chế độ ăn giới hạn. Heintz và cộng sự ức chế nhiều thành tố trong đường tín hiệu mTORC1 và quan sát vai trò của SFA-1 trong việc kéo dài tuổi thọ. Các thí nghiệm chỉ ra rằng vai trò của SFA-1 cao hơn RAGA-1. Nó như một chất điều tiết chung (general regulator) của các thành tố đường tín hiệu mTORC1, bao gồm các yếu tố AMPK, RAGA-1, và RSKS-1/S6 kinase.
Dựa trên các dữ liệu, Heintz và cộng sự đề xuất một cơ chế “thô” về vai trò của SFA-1 trong chế độ ăn hạn chế DR và đường tín hiệu mTORC1 (Hình 3). Mặc dù vậy, làm thế nào mà SFA-1 điều hòa sự trường thọ DR và mTORC1 vẫn còn chưa rõ ràng.

Hình 3: Mô hình vai trò của SFA-1 trong tính trường thọ (longevity) do DR và mTORC1.
Tuy nhiên, các kết quả của công trình này đóng vai trò quan trọng trong hiểu biết hiện tại về cắt tiền RNA thông tin trong trường thọ do chế độ ăn hạn chế và do ức chế mTORC1. Một hướng đi cho việc ứng dụng công trình này là tính tiềm tàng trong việc ức chế mTORC1 hoặc trong việc sử dụng chế độ ăn giới hạn để điều trị các bệnh liên quan đến cắt tiền RNA thông tin ở người.
Nguồn: http://ibsgacademic.com/2016-12-pre-mrna-splicing-longevity/






Không có nhận xét nào:
Đăng nhận xét